
By Steve Drury Firts PUBLISHED ON November 5, 2021
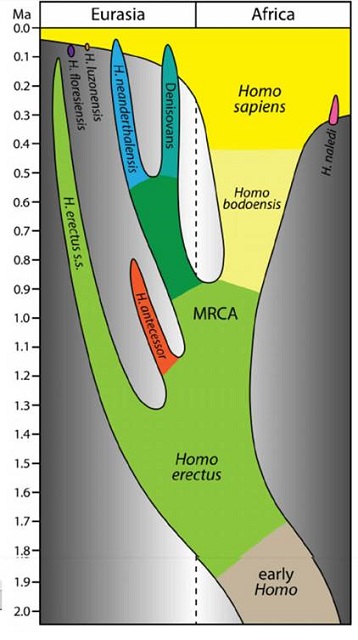
Palaeoanthropologists are in a bit of a muddle about the early humans of the Middle Pleistocene (~780 to 130 ka), namely Homo heidelbergensis and H. rhodesiensis.
The first was defined in 1907 based on a massive lower jaw or mandible (but no cranium) found near Heidelberg in Germany. Fourteen years later a massively browed cranium (but no mandible) turned up near Kabwe in what is now Zambia (then Northern Rhodesia). That specimen became, in true colonialist fashion, H. rhodesiensis.
Since then scientists have unearthed more such highly ‘robust’, ‘archaic’ remains in Africa, Asia and especially Europe: including at least 28 individuals in the Sima de los Huesos (‘pit of bones’), part of the World Heritage Site in the Atapuerca mountains of northern Spain.
Do these widespread fossils really represent just two species or do specimens just happen to fit within two broadly similar morphological types?
These days, most scientists experience discomfort with a reference to the legacy of Cecil Rhodes, so several sacks full of bones were metaphorically lumped into H. heidelbergensis. So widely dispersed are their sources and their ages covering such a wide span of time that the specimens might be expected contain a diverse range of genetic signatures. Yet only a single specimen from northern Spain, dated around 400 ka, has yielded DNA.
The Sierra de Atapuerca provided an even more archaic European dated between 1.2 to 0.8 Ma (Early Pleistocene), from which dental proteins have been extracted. Comparative proteomics have encouraged H. antecessor to be considered as a possible common ancestor for anatomically modern humans (AMH), Neanderthals and Denisovans … and H. heidelbergensis.

A group of palaeoanthropologists has proposed a way to clear such muddy waters (Roksandic, M. & Radović, P. et al. 2021. Resolving the “muddle in the middle”: The case for Homo bodoensis sp. nov.. Evolutionary Anthropology, v. 30, early-release article 21929; DOI: 10.1002/evan.21929).
Their device is to abolish the two previous species and lump together many human remains from the Middle Pleistocene of Africa into a new species named after the Bodo site in the Awash Valley of Ethiopia. It was there that a human cranium bearing characteristics similar to all the African specimens was found in 1976. Originally it was allocated to H. heidelbergensis, but now the composite group of archaic Middle Pleistocene Africans is proposed to be assigned to H. bodoensis. This composite species is also reckoned by the authors to be the ancestor of all surviving, anatomically modern humans.
European examples of H. heidelbergensis are to be slotted into an early population of Neanderthals. Since the Denisovans of Asia are only known by DNA from tiny skeletal fragments, the taxonomic rearrangement logically should assign Asian archaic humans to early members of that mysterious but well-defined group. But a spanner in the works is that the sole example of H. heidelbergensis DNA (mitochondrial) – from northern Spain – more closely resembles Denisovans than it that of Neanderthals (see: Mitochondrial DNA from 400 thousand year old humans; Earth-logs December 2013).
There is also a bit of a problem with H. antecessor. There aren’t many specimens, and they are all from Atapuerca. Yet they are a plausible candidate, according to the proteomic analyses, for the most recent common ancestor (MRCA) of all subsequent humans (whatever taxonomists care to call them). But they do not fit in the taxonomic model suggested by Roksandic et al., who reject them as MRCA, on grounds that they are European. They consign them to an anomalous ‘spur’ that petred out in Spain while the real action was in Africa.
So what happens if a cranium that bears close similarity to both H antecessor and H. bodoensis pops out of African Early Pleistocene sediments (older than about 700 ka)? There is at least one candidate from ~1 Ma sediments in Eritrea (Abbate, E. and 16 others 1998. A one-million-year-old Homo cranium from the Danakil (Afar) Depression of Eritrea. Nature, v. 393, p. 458-460; DOI: 10.1038/30954), which is said to display ‘a mixture of characters typical of H. erectus and H. sapiens’. And there are others of that antiquity from Ethiopia.
Since the time of Charles Darwin there have been taxonomists who were (and are) either habitual ‘lumpers’ or ‘splitters’. There are more with a propensity for splitting because a new species carries the name of its initiator into posterity! So I expect the paper by Roksandic et al. to raise a cloud of academic dust. Yet taxonomic lumping has its stand-out species in the field of human evolution – H. erectus.
A great many ‘archaic-looking’ human remains from the period after ~1.9 Ma until as recently as 200 ka have been dubbed ‘Erects’, giving the group an unsurpassed survival span of over a million years. A few early examples from Africa have been ‘split’ away to give H. ergaster, on taxonomic grounds that some palaeoanthropologists do not fully accept. Yet there are signs of later diversity that ‘splitters’ have, so far, not dared to slice-off from the mainstream consensus. So common are these ‘Erect’ fossils in China, that it is almost state policy that it was they who gave rise to living Han Chinese people!
The lumpers are likely to hold sway in the absence of ancient DNA sequencing, which may never be possible outside temperate climates or for ages greater than that of the Spanish H. antecessor. With the knowledge that several anatomically very distinct hominin groups occupied the Earth together at several times in the last 300 ka – think H. floresiensis and H. naledi – it seems likely that the proposed pan-African H. bodoensis may not reflect past reality and the hypothesis needs considerably more testing.
If you’d like to read more of Steve’s blog…… https://earthlogs.org/homepage/
Many thanks to Steve Drury for permission to republish his article and to Bernie Bell for sending it into The Orkney News




Leave a Reply